










「一瞬、一瞬がドラマ、楽しかった私の大学生活60年!」 ・講師:河盛 隆造 先生 順天堂大学 名誉教授・日時:2026年5月14…

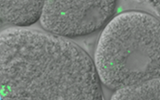
2026年3月13日, 東北大学加齢医学研究所の谷春菜助教をお招きし, 「ミトコンドリアゲノム変異がもたらす病態機構の理解と制御 –…

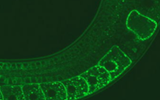
「オートファジーとリソソーム」 ・講師:内山 安男 先生 順天堂大学医学研究科・日時:2026年3月12日(木)16:00~・開催場…

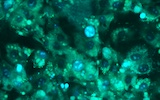
2026年3月4日、福岡大学理学部の小柴琢己教授をお招きし、「ミトコンドリアのストレス応答における分子基盤」というタイトルでご…

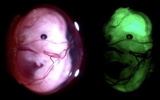
「Stem cell–derived beta cells for diabetes therapy: Do they engraft better than primary islets?」 ・講師:小松 弘武 …











![]()